
We have always been fascinated by what it means to be alive. The creation myths, among the oldest tales, talk about the source and nature of life. Mary Shelley's Frankenstein, often regarded as the first science-fiction novel, featured a central character obsessed with the quest to create life. As listeners, readers, and writers we have wondered what it really means for something to be alive, whether it's a bacterium, a computer program, or a planet. Scientists, writers, philosophers, and theologians have battled with the question. Competing definitions of life have come and gone. And still no one is much the wiser about what this thing called life really is.
Now, though, scientists are starting to follow Frankenstein's example. They are trying to understand the fundamental nature of life by creating it from non-life on a microscopic scale. Some stitch together pieces of various micro-organisms into a synthetic cell, while others mix simple chemicals trying to find the mysterious élan vital that will produce life. For all, though, the quest is to find what it means for something to be alive, to break down the wall between chemistry and biology.
However, these synthetic organisms don't just offer abstract scientific knowledge. Being wholly artificial, they could be specially designed for pollution detection and remediation, medical applications, hydrogen-energy production, pharmaceutical and plastics production, and more. As their genomes and even their physical structures are known and controlled precisely many environmental or medical problems could be designed out.
What is life?
So, if scientists are trying to make living things for these various uses, it would help if we could decide what it means for something to be alive. One definition says that living things have the capacity to absorb substances from their environment and hence to grow—but that can describe the growth of crystals. Or we might say that living things store energy and use it for purposeful activity—but that would also cover a wristwatch. Or we might say that living things produce offspring—but that might describe clouds which twist and divide and rain.
The current favorite description of life can be stated as "living things evolve." In other words, if something is to qualify as being alive, it must reproduce to give rise to offspring upon which natural selection can act.
Natural selection is the process by which differences between similar organisms cause the organisms to have different degrees of success at living. For example, one bacterial cell might be better at acquiring nutrients than another, or more efficient at using the nutrients that it does acquire, or better at surviving starvation. When nutrients are limited, these variants would gain an advantage and hence be more likely to survive and produce offspring.
If the search for life, then, is the search for things that undergo natural selection, how do we recognize the goal of our search? Three elements seem to be necessary for this: heredity, metabolism, and a container.
Heredity is information transmitted between generations. This is what makes offspring resemble their parents—sheep have lambs, and acorns grow into oak trees. Metabolism means any process on which selection can act. Any activity will do, as long as the organism's survival is affected by it. Examples include things like eating, breathing, or walking. Finally, the container is needed to hold the other two components together for long enough for selection to take place. Without a container, there's no "organism" because the information and metabolism aren't linked to one another.
Heredity itself must have two complementary characteristics if natural selection is to occur: offspring must resemble their parents for the successful characteristics to be passed on to subsequent generations; but offspring must not resemble their parents too closely. If offspring are identical to their parents, there will be no differences for selection to work on and all offspring will have an equal chance of surviving.
This means that the machinery of inheritance must be error-prone, to produce offspring with unique characteristics. However, it must not be too error prone or variation will swamp the accurate transmission of information between generations. An offspring's survival would not be affected by how successful its parent was, because its own characteristics would be too different.
In natural life, the molecule that controls heredity is a nucleic acid, usually DNA. This stores information as the sequence of nucleotide bases along its linear molecule; these are our genes. The collection of genes in an organism is known as its genome. The metabolism of a cell describes all of its activities, such as feeding or movement. These activities are mostly carried out by proteins. So, there needs to be some communication between the DNA and the protein, so that the proteins know what to do and when. This communication function is started by another nucleic acid, RNA. Thus, nucleic acids store and transmit information, whereas proteins do the work.
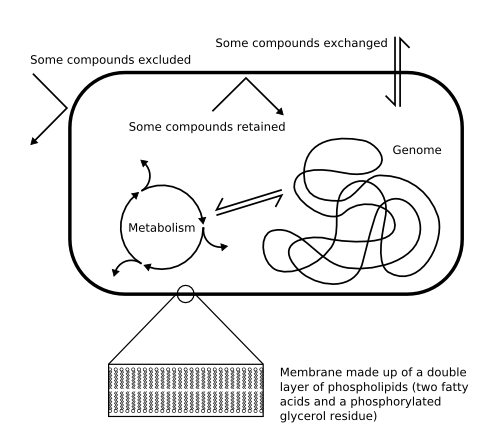
Figure 1: A conventional, natural living cell;
Click to enlarge.
The container function in natural life is provided by a cell membrane. This is a double layer of fat molecules that surrounds the watery cell contents and holds them together, dividing the water inside the cell from the water outside. These fat molecules are called lipids, and the double layer structure is called a bilayer. As well as separating the inside from the outside, the cell membrane also allows certain substances to pass from one to the other in a controlled fashion (Fig. 1).
Minimal cell project
If this, then, is how natural living things are built, how are the new Frankensteins trying to emulate nature? There are three general approaches being taken. The first is to stick very closely to nature, and to try and copy what nature has done. The second is to be inspired by nature but to simplify things as far as possible so that we can understand them better. The third is to start from scratch and to design something totally new.
Our first group trying to design an organism is taking the most straightforward approach—start with a normal cell and replace its genome with a synthetic one. J. Craig Venter and Hamilton O. Smith from the Venter Institute (Rockville, MD) are the leaders of the Minimal Cell Project. Venter is famous from his involvement in the sequencing of the human genome; indeed, he recently became the first individual to have his own personal genome completely sequenced. That he is putting his weight behind the Minimal Cell Project means that he believes that it could succeed.
And this approach is indeed the one that is most likely to work, and soonest. The Venter Institute's method is based on the idea of a "minimal genome"—a genome that contains the relatively small set of genes that are expected to be the only ones required to keep an organism alive. For example, a bacterium such as E. coli has around four-and-a-half million base pairs, encoding about 4000 genes. However, based on analyses of the genomes of some very thrifty bacteria, it is thought that a minimal genome might require only 200 or so genes. The other genes encode functions that a simple minimal cell would not need, such as finding food or responding to environmental changes.
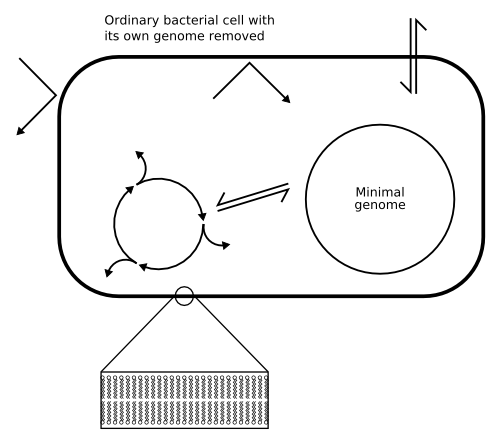
Figure 2: The minimal cell;
Click to enlarge.
Once the team have decided exactly what their minimal set of genes is, they will synthesize the minimal genome in the lab, insert it into a bacterium whose own genome has been removed, and see whether the resultant cell can grow and reproduce (Fig. 2). Their problem, though, is that current artificial DNA synthesis methods lag far behind the natural ones as only short stretches of DNA can be synthesized. Making an entire genome of tens of thousands of bases will be much more difficult than this, but Venter believes that it could be complete within the next few months.
As a first step towards this goal, the team synthesized the entire 5386-base genome of the phi X 174 virus in 2003. They did this by joining short DNA strands together in sequence to make the whole virus genome. The complete synthesis took just 14 days. A bacterial genome, even a minimal one, would be exponentially harder than this because of its much greater size. But, nonetheless, this demonstrates that it is possible in principle to build an entire genome in the laboratory.
In June 2007, at the Synthetic Biology 3.0 conference in Zurich, Switzerland, the team also reported that they had managed to replace the genome of one bacterium with the genome of another. In effect, they have managed to do a genome transplant. It would only be a short step to transplant their minimal genome into a bacterium that has had its own genome removed. This would create a hybrid cell that uses the bacterial cell to kick-start the genome into life.
With these successes under their belts the Venter team are the favorites to reach their goal first, perhaps within the next year. (On October 20, 2007, New Scientist magazine published an interview with Craig Venter in which he said that his team were in the process of publishing their successful construction of their minimal genome, and that we should expect the working synthetic organism "relatively soon.") It will be a significant achievement to produce a synthetic organism this way. However, it won't necessarily tell us very much about what life is in general, as the organism is kick-started using an intact bacterial cell. That is, it gains intact versions of many proteins and structures from its host and doesn't need to create them itself. Thus, although the final product is synthetic, it will still be "life as we know it" and won't help us to come to firm conclusions about what life really is.
Artificial cells
Taking one step beyond this approach, Pier Luigi Luisi and his group at the University of Rome 3, Italy, are also planning to use a synthetic minimal DNA genome, like the Venter Institute. However, instead of using a pre-existing bacterial cell, they plan to create their own, building a new synthetic cell from the ground up using readily available or synthesizable chemicals, and to add in components until the cell starts to work—and hence comes to life.
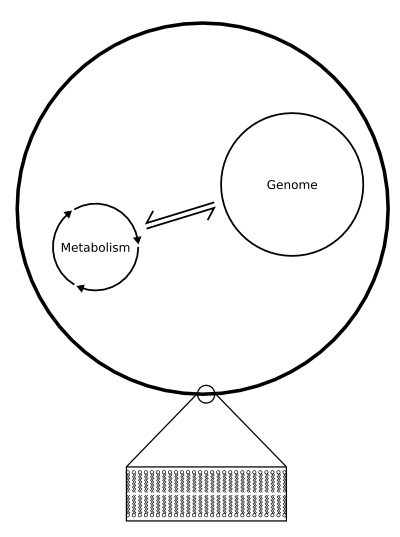
Figure 3: An artificial DNA-based cell;
Click to enlarge.
First, they make a cell membrane by synthesizing liposomes. These are spherical lipid bilayers like the cell membranes of normal cells, but created artificially from known lipid molecules. They then add to these liposomes the components they find necessary for their artificial cells to become living organisms, such as enzymes and other proteins. Eventually, the hope is to create a system that works like a simple bacterial cell, except that it will have been created from scratch (Fig. 3).
Luisi's group has already managed to enclose DNA and RNA molecules within liposomes, and to persuade this RNA and DNA to replicate within the liposomes. In June 2007, at the Synthetic Biology 3.0 conference, the group reported that they had successfully managed to make functional protein in a liposome "cell." The major obstacle is combining all these activities into a single system.
With metabolism dealt with, these liposomes face another hurdle when it comes to being alive—they have to reproduce. That is, one liposome has to split to form two smaller liposomes as offspring. In normal cells, a whole array of molecules is called into play to make cells divide, and to ensure that each daughter cell contains everything that it needs to survive. However, in a basic artificial cell, this will hopefully be unnecessary. For example, we might not care initially whether all daughter cells inherit a genome, as long as some do. If nothing else, this will give natural selection something to work on!
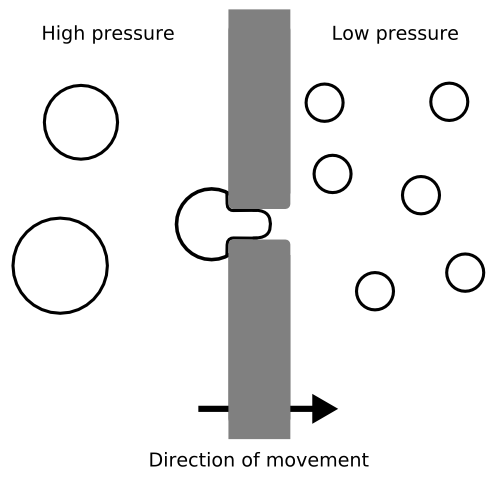
Figure 4: Propagating artificial cells using simple mechanical force and a filter;
Click to enlarge.
To make their liposomes divide, Luisi's group has at the moment turned to a simple system that pushes the liposomes through holes in a filter (Fig. 4). This causes large liposomes to split into smaller ones as they emerge from a hole. All the progeny tend to peel off at the same size, so a parent liposome that has grown larger than this will be divided into several offspring.
It should be possible to generate a population of liposomes that grow using their genome and metabolism, then to pass them through a filter so that they divide, and finally to return the offspring to the start so that they can grow once more. This would allow an artificial cell that lacks the machinery to divide by itself to propagate and be acted on by natural selection—and so potentially to be alive.
This project, when it bears fruit, should tell us more about what life really is because it will have examined the essential elements far more closely, in order to determine which to include within the synthetic cell. However, we are still considering only the conventional DNA-RNA-protein cell of earthly life. Moving beyond this requires a leap of faith to find other molecules that can carry out the tasks of life.
Leaving DNA behind
Jack Szostak (Department of Molecular Biology, Massachusetts General Hospital) is facing this challenge and discarding DNA from his artificial cells. Instead, Szostak is turning the clock back to a world that many biologists believe came before DNA. When life first arose, it is thought that the original molecule that carried the information of heredity was not DNA but its poor cousin RNA.
RNA is less stable than DNA and doesn't form the neat double-helix structure that we are all aware of from school biology lessons. However, life based on RNA has two big advantages over that based on DNA. First, it is much simpler—instead of a three-tiered structure with DNA storing information, RNA transmitting information and protein doing the work, there would be only two (RNA storing information and protein doing the work).
Second, and even more important, RNA has a trick that DNA doesn't—it is chemically active and can act as an enzyme. RNA enzymes are called ribozymes and many natural examples are already known. They persist in our cells even today, carrying out some essential functions. Indeed, the basic function of turning information from DNA into protein involves a large, complex structure called the ribosome, which includes both protein and RNA components. This means that a synthetic cell might even be reduced to one level—RNA both storing information and doing the work.
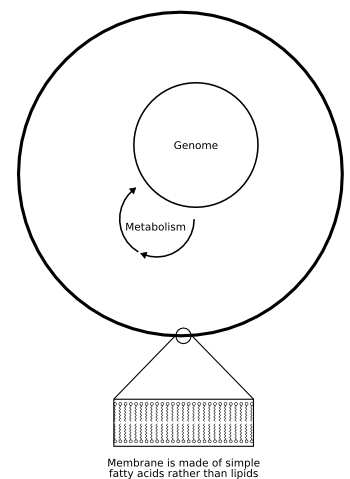
Figure 5: A simple RNA-based cell;
Click to enlarge.
So, Szostak's team are using liposomes similar to those being studied by Luisi's lab to create synthetic cells, but with much simpler contents (Fig. 5). First, they are using a self-replicating boundary to their cell, made of simple fatty acids. This is "self-replicating" because, unlike lipid molecules, free fatty acids can dissolve in the cell's environment and then spontaneously incorporate themselves into the membrane upon contact with it.
Second, they want a self-replicating genome. That is, they want an RNA molecule that catalyses its own formation—an RNA molecule that is an RNA polymerase. To produce this, the group are using directed molecular evolution to generate improved RNA-polymerase ribozymes. Some have already been produced but they tend to be inefficient and to be fairly inaccurate at copying RNA.
This inefficiency of ribozymes is believed to have been one of the driving forces that led to the use of protein in cells—proteins are more structurally complex than RNA and DNA, and hence offer a wider range of activities and efficiencies. Despite this limitation, Szostak's team hope that their ribozymes will be "good enough."
Third, they need a mechanism to allow changes in the genome to affect the performance of the cell and hence to allow natural selection to act. That is, they need a metabolism for their cell. There are several possible ways they might generate this. For example, the cell could include an RNA molecule that has another enzymatic activity, perhaps synthesizing fatty acids that would then help the cell's membrane to grow. This second RNA would be copied by the ribozyme genome in the same way that the genome copies itself. Alternatively, the cell could include an RNA that alters the structure of the cell or its rate of growth.
Another possibility, which Szostak and colleagues published in the journal Science three years ago, is to use simple osmotic pressure (the tendency of water to enter or leave the cell) as the basis of selection. When RNA molecules are trapped within liposomes, they affect the osmotic balance of the cell in a way that affects cell growth—cells containing RNA grow at the expense of empty cells or cells with less RNA. When the fatty acids needed for growth are limiting, there is thus competition for the scarce resource, allowing selection to act to encourage rapid RNA replication, leading to increased RNA content, and hence faster growth. Indeed, the less-efficient cells actually shrink and "die" when out-competed, which suggests that natural selection would be strong in such a system. One great advantage of this approach is that it effectively combines selection and inheritance, in that the molecule that acts as the genome also produces a direct effect that influences growth and survival.
This approach to creating synthetic cells takes us rather further away from the earthly life that we know and should give some real insights into what makes a system move from non-living to living. Indeed, these systems are often talked about more in terms of chemistry than biology; their simplicity is a far cry from the complex and sophisticated machinery of even the simplest bacterium. However, it does this partly by looking back into our evolutionary history and so might not truly broaden the horizons of what we think of as alive. After all, however simple it is, it is still based on the same molecules and structures that we find in our own cells.
Beyond RNA
In the Los Alamos laboratory in New Mexico, USA, a team led by Steen Rasmussen is reaching into the realms of fantasy as they design their own synthetic life form. In their quest for simplicity, they have thrown away many of the assumptions that we make about living things and ended up with a creature that is perhaps more complicated than Szostak's simple RNA cell but is much stranger and possibly even more interesting.
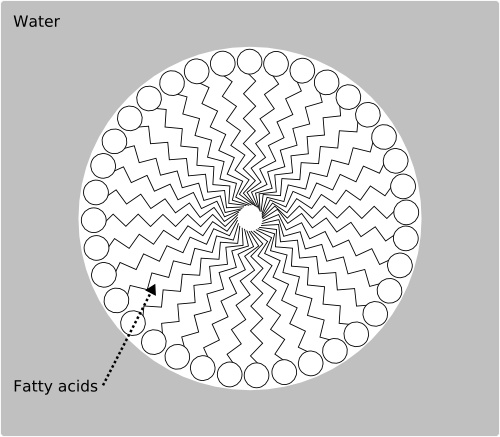
Figure 6: The structure of the Los Alamos bug;
Click to enlarge.
All life on Earth depends on water as its solvent. This is why it requires the lipid membrane to divide the water inside the cell from the water outside, and to keep the chemicals dissolved in that water close together. By contrast, Rasmussen's "Los Alamos bug" is oil based. Instead of a lipid bilayer bounding the cell, it is based on a simple micelle—a droplet of fatty acids suspended in water (Fig. 6). Thus, the container that is required for natural selection to occur is a simple consequence of the chemistry: oil and water don't mix. All of its components are oil soluble instead of water soluble and, as you might imagine, this makes a huge difference to what those components are.
With an oil-based organism, the hereditary material must also be rethought. Rasmussen's team has stuck with the idea of nucleic acids because of their known ability to carry information but, instead of using DNA or RNA, they have gone with a related compound—peptide nucleic acid (PNA). This molecule has the useful property of existing in two forms: a double PNA chain akin to the DNA double helix that is soluble only in oil; and lone PNA chains that are also attracted to water. The group hope to use this feature of the molecule as the basis of its heredity and also to allow selection to occur (Fig. 7).
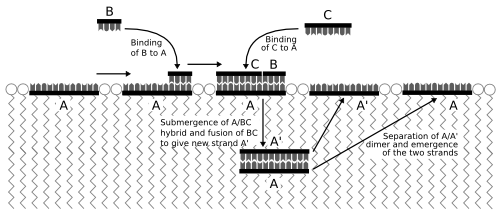
Figure 7: The cycle of PNA replication in the Los Alamos bug;
Click to enlarge.
The PNA genome of the bug replicates by acquiring small fragments supplied as its food. When in single-stranded form, the long genome molecules of the bug are exposed on the surface of the cell, with their nucleotide bases exposed to the environment. The "food" fragments stay in the water until they come into contact with a matching PNA genome sequence in a bug cell, where they bind to it. Once the long genome molecule is all bound to these small fragments, its structure changes to the double-stranded form. The molecule therefore now submerges within the oily interior of the cell. Inside the cell, the fragments are fused by photons of light into a new single genome strand. After a while, the two strands separate and emerge once more on the cell surface, ready to pair with new fragments and to continue to replicate the genome.
Unlike the genetic systems in the other synthetic organisms we've already encountered, the "genome" of the Los Alamos bug would not store any information in its sequence. That is, the bug won't have any actual genes. Instead, the team are going to rely on the fact that PNA molecules with different sequences behave slightly differently. This means that the PNA genome will affect the properties of the cell directly, without having to be converted into anything else. This will lead to selection for those PNA variants that give advantages in reproduction and survival.
The growth of the Los Alamos bug is also driven by light, analogous to the way plants and many bacteria use light to drive their own metabolisms. However, in the same way that the container and genome of the bug have been simplified as far as possible, so has its metabolism. The basic metabolic process is just growth—the inclusion of new fatty acid molecules into the cell. This causes its size to increase ready for reproduction, which is just the division of the cell to form two new cells.
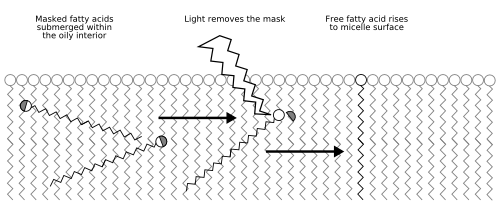
Figure 8: The metabolism of the Los Alamos bug;
Click to enlarge.
Fatty acid molecules usually dissolve in water but, for the bug's food, they are modified so that they are fat soluble. The molecules will therefore tend to gather in the interior of the oily micelles that make up the bug's cell. Light is then used to remove the mask, restoring it to a normal fatty acid. This will then take up a position in the cell's structure with its water-loving end outermost and its fatty tail pointing in (Fig. 8). Again, this process could be affected by the sequence of the PNA genome, because PNA can transfer electrons from one place to another, and this can drastically affect photochemical reactions. Thus, there is another way for natural selection to operate and for genetic information to accumulate.
So, the Los Alamos bug has a life cycle that runs like this: From the time it is first "born," the cell starts acquiring fatty acids from the surrounding solution and converting them into cell components, and hence growing. Meanwhile, the PNA genome is acquiring template fragments from the external environment. Exposing the cell to light makes the PNA strands join together, and also unmasks the fatty acids so that they emerge and increase the cell's surface area.
The structure of the spherical cell becomes unstable once it reaches a certain size—there are too many water-loving fatty-acid molecules trying to reach the water for a spherical shape to hold them all. The micelle then splits into two, each part carrying away some of the copies of its parent's genome, ready to start again—a primitive living organism.
Shadow of Frankenstein
The Los Alamos bug promises us new and startling insights into what really makes a living organism. Other workers are trying even stranger chemistries in their attempts to create life. Doron Lancet (Weizmann Institute of Science, Israel) is studying a system composed only of lipids, with no DNA or other nucleic acids and no protein. By studying groups of lipid molecules that catalyze their own assembly, Lancet believes that it will be possible to create extremely simple organisms.
By contrast, Antón Vila-Sanjurjo (UC Berkeley, California, USA) is trying to reverse the ancient endosymbiosis between animal cells and their mitochondria, in order to create a minimal cell like that envisioned by the Venter Institute team. Mitochondria have such a small genome that there is almost nothing there already to interfere, and they have a ready-made life-support system already available in their host cells.
Whichever approach succeeds first and best, this work will answer some crucial questions—most particularly, how life really can emerge spontaneously from chemistry. This alone will be a remarkable demonstration of our own likely origins.
Of course, all these systems require external inputs if they are to survive. The Los Alamos Bug needs PNA fragments and light pulses; the liposome-based systems require physical intervention if they are to replicate. However, all living things require external inputs of nutrients and energy. We ourselves need oxygen from air, food from plants or animals, and water. Synthetic life forms, though, will be particularly fragile and fussy.
And this provides a crucial safeguard against one of the eternal worries of biotechnology—will we create a Frankenstein's monster who will turn and kill us? In Mary Shelley's story, Frankenstein was usurping the role of his own Creator by seeking to make life, and was repaid for his hubris. Just so, are we on dangerous ground somehow by invading what some would claim as God's sole preserve? Let us hope not. Perhaps here is where we start using Nature's tools to continue the long experiment of life, in new and previously unimagined directions.
Frankenstein's sin was lack of care for his creation. Provided that we take the proper precautions, there is no reason to fear these new forms of life. Indeed, they promise us a deeper understanding of ourselves and even of our place in the Universe. For if we find that life is easy to create in the lab, that suggests that it is more likely to arise naturally on other planets. If we find that radically different chemistries and structures can form living systems, that broadens the range of things we must look for when we search for life among the stars. Rather than "what ifs," we will have laboratory evidence, and a clearer understanding of what divides chemistry from biology.
What does seem certain is that, as the science continues and we start to understand more about what distinguishes living things from non-living matter, we will be providing plenty of material for scientists, philosophers and theologians to debate. Long before any of these efforts to create synthetic life bear fruit, this debate should have started to give us more insight into the fundamental nature of life, and perhaps even into how life itself got started here on Earth.
Further reading
Smith HO, Hutchison CA 3rd, Pfannkoch C, Venter JC (2003) Generating a synthetic genome by whole genome assembly: phiX174 bacteriophage from synthetic oligonucleotides. Proceedings of the National Academy of Sciences of the USA 100, 15440-15445
Walde P (2000) Enzymatic reactions in giant vesicles. In: Giant Vesicles, Perspectives in Supramolecular Chemistry (Luisi P, Walde P, eds), pp. 297-311, John Wiley & Sons
Chen IA, Roberts RW, Szostak JW (2004) The emergence of competition between model protocells. Science 305, 1474-1476
Rasmussen S, Chen L, Stadler BMR, Stadler PF (2004) Proto-organism kinetics: evolutionary dynamics of lipid aggregates with genes and metabolism. [Editor's note: PostScript file; converted to text] Origins of Life and Evolution of the Biosphere 34, 171-180